Bioquímica Actividad Antihiperglicemiante de Bauhinia megalandra.(Revisión)
Efectos de B. megalandra sobre la glucogenólisis hepática
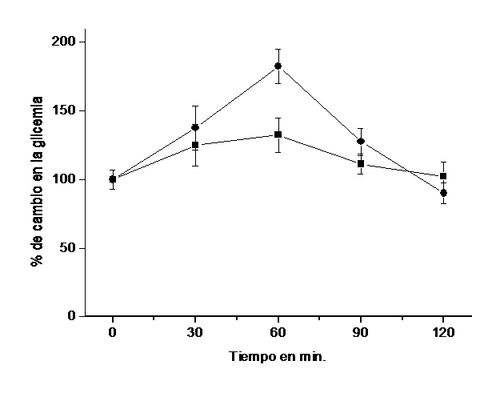
Como podemos observar en la Figura 7, al inyectar adrenalina
intraperitonealmente (13), se incrementó la glicemia con un máximo
de 82 %a los 60 min. La administración
oral del extracto de las hojas de B.
megalandra 60 min. antes de la inyección de la catecolamina, redujo en
aproximadamente un 50 % la elevación de la glicemia producida por el efecto de
la adrenalina (14).
Figura 7. Efectos del
extracto acuoso de las hojas de B.
megalandra sobre la liberación de glucosa a la sangre estimulada por
adrenalina. Ratas macho de la
cepa Sprague Dawley fueron anestesiadas ligeramente (pentobarbital sódico 30
mg/Kg de peso) antes de administrarle por sonda gástrica agua (control ●) ó
3,44 mg/kg de peso del extracto de la planta (■). Luego de 1 h. a los animales
se les inyectó por vía intraperitoneal 0,15 mg/Kg de peso de adrenalina y se
midió la glicemia (29) cada 30 min. por 2 h. El 100 % representa el valor
promedio de la glicemiade ratas
alimentadas (7,2 ± 0,5 mmol/L). Los valores representan el promedio ± la
desviación estándar de 5 " 6 experimentos y en cada uno de los cuales se uso 3
ratas. A los 60 min. la diferencia entre control y experimental fue
estadísticamente significativa a p < 0,001.
Estos resultados son una evidencia indirecta de
la absorción intestinal de alguno(os) compuesto(s) presentes en el extracto
vegetal los cuales al alcanzar el hígado disminuyen la glucogenólisis por lo
cual no se eleva la glicemia por efecto de la adrenalina. La presencia de
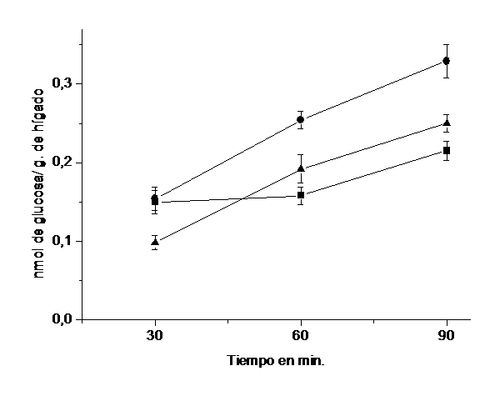
adrenalina en el medio de incubación de rebanadas de hígado de ratas
alimentadas, incrementa la liberación de glucosa en un 60 % y un 53 % a los 60
y 90 min de estudio respectivamente (Figura 8).
Figura 8. Efectos de la
adrenalina y del extracto de las hoja de B.
megalandra sobre la glucogenólisis hepática. Rebanadas de hígado de ratas fueron
incubadas en amortiguador Krebs-Ringer bicarbonato a pH 7,4 en atmosfera de O2/CO2
95/5 %, sin adición alguna para el control (■), o en la presencia de 2,4 ng de
adrenalina (●), o la combinación de 2,4 ng de adrenalina y 13,8 mg del extracto
de las hojas de B. megalandra (▲). A los tiempos indicados se determinó
la glucosa en el medio de incubación por el método de glucosa oxidasa-peroxidasa
(29). Los valores representan el promedio de 5 " 8 experimentos ± la desviación
estándar en cada uno de los cuales se utilizo 3 ratas. Las diferencias, a los
60 y 90 min., entre los valores
obtenidos con adrenalina y los encontrados en los controles y con la mezcla,
fueron estadísticamente significativos a p< 0,001.
Por el contrario la liberación
de glucosa por las rebanadas de hígado en la presencia simultánea de adrenalina
y del extracto vegetal, fue paralela y siempre significativamente menor a la
producción de glucosa en presencia de adrenalina sola, mostrando 38; 25 y 24 %
de inhibición a los 30; 60 y 90 min. respectivamente. Así mismo a los 30 min.
de incubación, la presencia simultanea de adrenalina y el extracto de la planta
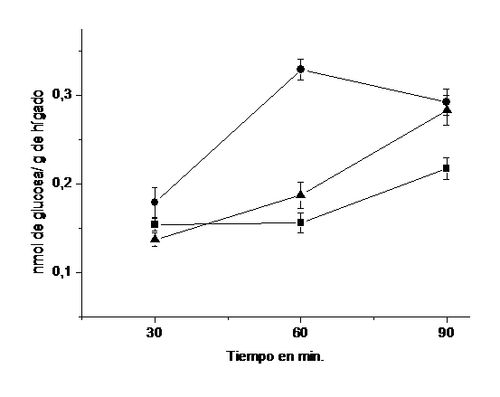
reduce en un 38 % la producción de glucosa en relación al control (14).En la Figura 9 se muestra el efecto del dibutiril-AMPc
sobre la producción de glucosa por rebanadas de hígado de rata (14);
como puede observarse la presencia del nucleótido condicionó un incremento en
la liberación de glucosa por las rebanadas de hígado a todos los tiempos
estudiados, con un máximo a los 60 min.
Figura 9. Efectos del
dibutiril AMPc y del extracto de las hojas de B. megalandra sobre la glucogenólisis
hepática. Rebanadas
de hígado de ratas fueron incubadas en amortiguador Krebs-Ringer bicarbonato a
pH 7,4 en atmosfera de O2/CO2 95/5 %, sin adición alguna
para el control (■), o en la presencia de 0,01 μM de dibutiril AMPc (●), o la
combinación de 0,01 μM de dibutiril AMPc más 13,8 mg del extracto de las hojas de
B. megalandra (▲). A los tiempos indicados se determinó la glucosa en el medio de
incubación por el método de glucosa oxidasa-peroxidasa (29). Los valores
representan el promedio de 5 " 8 experimentos ± la desviación estándar en cada
uno de los cuales se utilizo 3 ratas. Las diferencias, a los 60 min., entre los
valores obtenidos con dibutiril AMPc y los encontrados en los controles y con
la mezcla, fueron estadísticamente significativos a p< 0,001.
La presencia simultanea del extracto
acuoso de B. megalandra y dibutiril
AMPc provocó una disminución del efecto del nucleótido la cual fue más marcada
a los 60 min cuando se observó una reducción de un 43 %. Los resultados
obtenidos en estas condiciones ex vivo
(Figuras 8 y 9) evidencian que los compuestos presentes en las hojas de B. megalandra afectan un evento
posterior a la producción del AMPc, probablemente la actividad de la G-6-Pasa. La única reacción
que tienen en común las vías glucogenólisis y neoglucogénesis es la catalizada
por la enzima G-6Pasa. En consecuencia el conjunto de resultados, presentados
hasta ahora, sugieren fuertemente que los flavonoides presentes en las hojas de
B. megalandra inhiben la G-6-Pasa lo cual se traduce en
una reducción de la capacidad de aportar glucosa a la sangre por la glucogenólisis
y la neoglucogénesis hepática. Dichos flavonoides pudieran ser útiles en el
tratamiento de la diabetes no insulino dependiente ya que muestran un efecto
antihiperglicemiante.
Efectos de B.
megalandra sobre la absorción intestinal de glucosa.
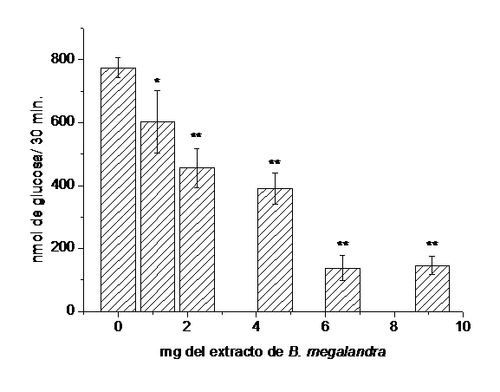
Usando como modelo experimental segmentos intestinales aislados in situ, demostramos (15) que
la absorción intestinal de glucosa es inhibida por el extracto acuoso de las
hojas de la planta en una manera dependiente de la dosis (ver Figura 10).
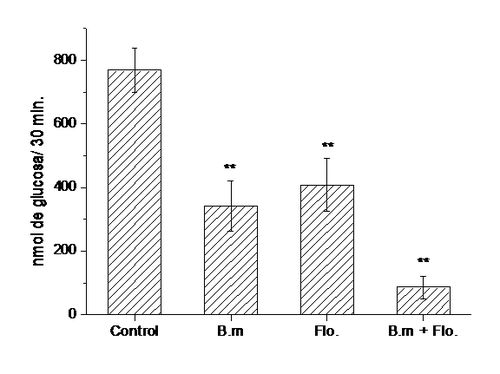
Figura 10.
Efectos del extracto de B. megalandra sobre
la absorción intestinal de glucosa. En segmentos intestinales in situ se inyecto 1 mL de 10 mM glucosa, 0,9 % NaCl solamente (0) o con
la adición de cantidades crecientes del extracto de de las hojas de B. megalandra (1,14; 2,28; 6,5 y 9,1
mg).A los 30 min se recuperó el líquido
remanente y se cuantificó la glucosa absorbida utilizando el método de glucosa
oxidasa-peroxidasa (29). Los valores corresponden al promedio de 8 experimentos
± la desviación estándar. * significa p<0,005 y ** significa p< 0,0005 de
acuerdo a la prueba t de Student.
Al
estar presente simultáneamenteel extracto
acuoso de B. megalandra y floricina,
un conocido inhibidor de SGLT 1 (16),se observó un efecto inhibitorio aditivo lo
cual sugiere que en la planta están presente compuestos capaces de inhibir
dicho transportador (ver Figura 11).
Figura 11. Efectos del extracto de B. megalandra y la floricina sobre la absorción intestinal de
glucosa. En segmentos intestinales in situ se inyecto 1 mL de 10 mM glucosa, 0,9 % NaCl solamente
(Control) o con la adición de 4,55 mg del extracto de de las hojas de B. megalandra (B.m) ó 0,1 mM floricina (Flo) ó
4,55 mg B.m. + 0,1 mM
floricina (B.M. + Flo).A los 30 min se
recuperó el líquido remanente y se cuantificó la glucosa absorbida utilizando
el método de glucosa oxidasa-peroxidasa (29). Los valores corresponden al
promedio de 8 experimentos ± la desviación estándar. ** significa p< 0,0005
de acuerdo a la prueba t de Student.
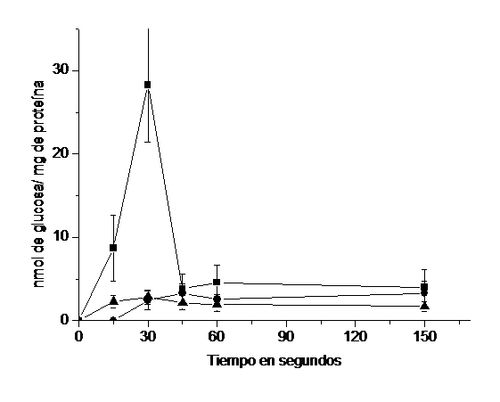
Al
incubar vesículas de membrana apical de enterocitos en un medio externo con 14C-glucosa
y Na+ comprobamos el típico comportamiento con un pico máximo de
captación de glucosa a los 30 seg. para luego decaer (17). En
presencia del extracto de la planta o de floricina se elimina el pico de
captación, resultado que claramente indica que en el extracto de la planta
existen compuestos capaces de inhibir el transportador SGLT 1, confirmando lo sugerido de acuerdo al experimento anterior
(15). La inhibición de la absorción intestinal de glucosa por el
extracto de ésta planta, afectando al SGLT 1, es similar al reportado para los
polifenoles del te verde (18) y las isoflavonas de soya (19)
pero diferente al ejercido por la acarbosa, la cual inhibe la digestión de los
carbohidratos y no la absorción de los mismos (20). La
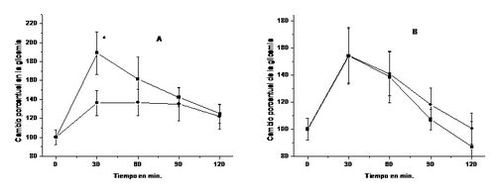
administración simultánea, por vía oral, de glucosa y del extracto foliar
condiciona una curva de tolerancia glucosada casi plana (Figura 13).
Figura 12.
Efectos del extracto acuoso de las hojas de B.
megalandra y de la floricina, sobre las captación de 14C-glucosa
por vesículas de borde apical de enterocito. Se midió la captaciónde 14 C-glucosa por vesículas
de membranaapical de entericito (17)
por el método de filtración rápida (29) en la ausencia de adición alguna (-■-)
o de la presencia de 4,55 mg del extracto de las hojas de B. megalandra (-▲-) o de 1 mM floricina (-●-). Los resultados se
expresan como nmol de glucosa captados/ mg de proteínas de membrana y
representan el promedio ± la desviación estándar de 9 experimentos en los
cuales se utilizó al menos 2 ratas.
Figura
13. Efectos del extracto acuoso de las hojas de B. megalandra sobre la tolerancia glucosada. Las ratas recibieron 1 g de glucosa/ Kg de peso por
vía oral (A) o por inyección subcutánea (B) junto con 263 mg del extracto de la
planta (-●-) o un volumen equivalente de agua (-■-) por vía oral. A los tiempos
indicados se determinó la glicemia (29), la cual al tiempo cero fue 4,17 ± 0,52 mM; los valores se
expresan en términos porcentuales del mismo y corresponden al promedio de siete
experimentos ± la desviación estándar. * diferencia estadísticamente
significativa a p< 0,005.
Por el contrario, cuando la glucosa se
administró subcutáneamente y el extracto vegetal por vía oral no se observó
efecto alguno sobre la curva de tolerancia glucosada. Estos resultados muestran
claramente que el extracto de la planta afecta la absorción intestinal de
glucosa y no otros factores relacionados con la glicemia tales como la
producción y/o liberación de insulina ni el consumo de glucosa por los tejidos (15).
Estudiamos el efecto que los flavonoides: kanferol 3-O-α-ramnosa, quercetina
3-O-α-ramnosa y kanferol 3-O-α-(2"
galoil)-ramnosa, purificados antes por nosotros (10), ejercen sobre
la absorción intestinal de glucosa, encontramos que solo el kanferol
3-O-α-ramnosa disminuyó de una manera apreciable (28 %), la absorción
intestinal de glucosa (Figura 14). En relación a la estructura química y la
actividad biológica vale la pena destacar: a) la presencia del grupo galoilo
unido a la ramnosa anula la capacidad inhibitoria del kanferol 3-O-α-ramnosa
sobre la absorción intestinal de glucosa; b) la ramnosa no es determinante para
condicionar la actividad biológica, ya que esta presente en ambos flavonoides y
solo uno de ellos inhibe la absorción intestinal de glucosa y c) la presencia
de un H (kanferol) en lugar de un OH (quercetina) en la posición 3" del anillo B del flavonoide,
es determinante para condicionar la inhibición de la absorción intestinal de
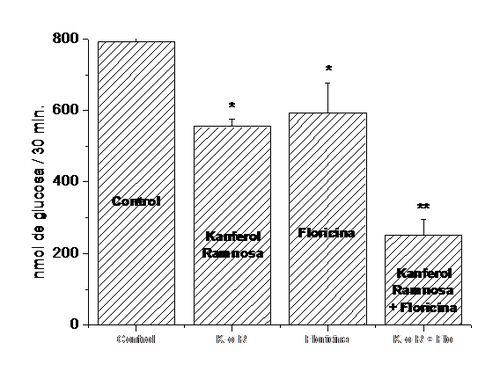
glucosa. (21)Como se muestra
en la Figura
14, la presencia conjunta de kanferol 3-O-α-ramnosa y floricina inhiben la
absorción intestinal de glucosa (68 %) en más del doble del efecto de
cualquiera de los dos compuestos solos (kanferol 3-O-α-ramnosa 28 % y floricina
25 %).
Figura 14. Efectos del kanferol
3-O-α-ramnosa y de la floricina sobre la absorción intestinal de glucosa. En
segmentos intestinales aislados in situ, se
inyecto 1 mL de NaCl 0,9 %, 10
mM glucosa y DMSO 2 % para los controles y en los
experimentales con la adición de: kanferol 3-O-α-ramnosa 5 mM o floricina 0,1 mM o kanferol
3-O-α-ramnosa 5 mM
+ floricina 0,1 mM.
Después de 30 min la glucosa remanente en los segmentos intestinales fue medida
por el método deglucosa oxidasa-peroxidasa
(29) y la cantidad absorbida fue calculada por diferencia. Los valores
corresponden al promedio ± la desviación estándar de 3 experimentos. *
significa p< 0,05; ** significa p< 0,001 de acuerdo al prueba t de
Student.
Estos resultados son una clara evidencia del efecto sinérgico del
kanferol 3-O-α-ramnosa y la floricina, un inhibidor conocido del SGLT 1 (16).
En la Tabla 5
se muestran los valores cinéticos: KM y VMAX, de la
absorción intestinal de glucosa en ausencia, control, y presencia de kanferol
3-O-α-ramnosa. Los valores controles son diferentes a los reportados por Ader y
Col. (22) y por Li y Col. (23), al respecto es importante
destacar que en ambos trabajos los parámetros cinéticos publicados corresponden
a los del SGLT1 y en nuestro caso conciernen a todo el proceso de la absorción
intestinal de glucosa. El kanferol 3-O-α-ramnosa se comporta como un inhibidor
competitivo de la absorción intestinal de glucosa ya que incrementa el KM con
una disminución de la VMAX
no significativa estadísticamente (21). De los resultados anteriores
podemos concluir que el kanferol 3-O-α-ramnosa es un inhibidor competitivo del
SGLT 1. Utilizando extracciones con solventes de polaridad creciente y
cromatografía en silica gel, purificamos parcialmente apigenina 8-glucósido, el
cual inhibió la absorción intestinal de glucosa en una manera sinérgica con la
floricina, lo cual sugiere que afecta al SGLT 1 (24)El uso de los flavonoides kanferol
3-O-α-ramnosa y apigenina 8-glúcosido pudiera
ser útil en el tratamiento de la diabetes tipo II ya que disminuiría el aporte
de glucosa a la sangre al reducir la absorción intestinal de misma,
comportándose como antihiperglicemiantes.
NOTA:Toda la información que se brinda en este artículo es de carácter investigativo y con fines académicos y de actualización para estudiantes y profesionales de la salud. En ningún caso es de carácter general ni sustituye el asesoramiento de un médico. Ante cualquier duda que pueda tener sobre su estado de salud, consulte con su médico o especialista.
Instituto de Medicina Tropical - Facultad de Medicina - Universidad Central de Venezuela.
Elaborado por el Centro de Análisis de Imágenes Biomédicas Computarizadas CAIBCO, caibco@ucv.ve
Este portal ha sido desarrollado gracias al apoyo del Fonacit