Monografías docentes
Glucagón y la hipótesis bihormonal de la diabetes
Regulación del metabolismo hepático de la glucosa por el glucagón
El efecto hiperglicemiante del glucagón fue una de las primeras actividades biológicas establecidas para ésta hormona y la misma se debe al incremento en la glucogenólisis y en la neoglucogénesis8. Como se discutió antes, la unión del glucagón a GcGR estimula a la proteína Gαs la cual activa a la adenilato ciclasa, que a partir de ATP produce AMPc y éste último activa a la PKA (Figura 4), la cual fosforila en restos de serina y treonina a varias enzimas, entre las que destacan: la glucógeno fosforilasa quinasa (GPK por sus siglas en inglés), la glucógeno sintasa (GS por sus siglas en inglés), la fosfofructoquinasa 2 (PFK2 por sus siglas en inglés) y la piruvato quinasa (PK por sus siglas en inglés)(Figura 5). La GPK a su vez fosforila y activa a la glucógeno fosforilasa (GP por sus siglas en inglés) la cual produce fosforolisis del glucógeno (glucogenólisis) dando glucosa-1-fosfato que se transforma en glucosa-6-fosfato (metabolito común con la neoglucogénesis), la cual es hidrolizado por la glucosa-6-fosfatasa (G-6-Pasa), enzima que cataliza la última reacción de la glucogenólisis y de la neoglucogénesis, dando glucosa que pasa a la sangre incrementando al glicemia. Por otro lado la fosforilación de la glucógeno sintasa, la inhibe condicionando reducción de la glucogenogénesis (Figura 5)34. El glucagón estimula la neoglucogénesis e inhibe la glicolisis por la modulación de enzimas claves de ambas vías metabólicas. Mediante la vía de la PKA- CREB-CRTC2 se incrementa la expresión de los genes de la PEPCK y de la G-6-Pasa (Figura 4) por lo cual se incrementa de manera importante la neoglucogénesis. Por otro lado, la PKA fosforila a la PFK2, la cual en esta condición se inhibe su actividad de quinasa y se estimula su función de fructosa 2,6 bifosfatofosfatasa (FBPasa 2 por sus siglas en inglés), trayendo como consecuencia una disminución de los niveles de fructosa 2,6 bifosfato (F2,6P2), metabolito que es un modulador alostérico negativo de la fructosa 1,6 bifosfatofosfatasa 1 (FBPasa 1 por sus siglas en inglés) con lo cual se estimula la neglucogénesis (Figura 5). Por otro lado los bajos niveles de F2,6P2 disminuye la actividad de la fosfofructoquinasa 1 (PFK1 por sus siglas en inglés) trayendo como consecuencia inhibición de la glicolisis. Así mismo la PKA fosforila e inactiva a la piruvato quinasa hepática, lo que contribuye a la disminución de la glicolisis (Figura 5)34. Es interesante mencionar que el ATP y las coenzimas reducidas (NADH+H+) requeridos para permitir la neoglucogénesis son aportados por el incremento de la oxidación de los ácidos grasos promovida por el glucagón (ver más adelante). En resumen, el incremento de la glucogenólisis y de la neoglucogénesis aunado a la disminución de la glucogenogénesis y de la glicolisis condiciona el efecto hiperglicemiante del glucagón. 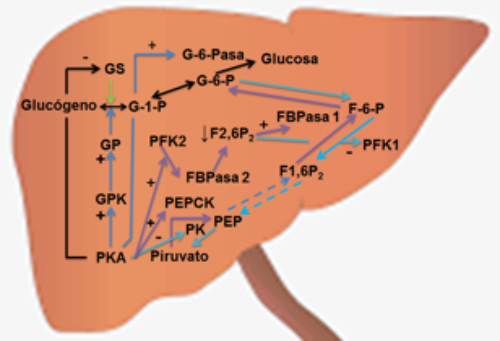
Figura 5. Regulación del metabolismo hepático de la glucosa por el glucagón.Como se destacó en la Figura 4, la unión del glucagón a GcGR condiciona la activación de la adenilato ciclasa la cual sintetiza AMPc, que activa a la proteína quinasa A (PKA) y ésta última fosforila varias enzimas; una de ellas la glucógeno fosforilasa quinasa (GPK) se activa y fosforila a la glucógeno fosforilasa (GP) la cual produce la fosforólisis del glucógeno dando glucosa-1-fosfato (G-1-P) que se transforma en glucosa-6-fosfato (G-6-P) la cual es hidrolizada por la glucosa-6-fosfatasa (G-6-Pasa) para dar glucosa libre que pasa a la sangre; la actividad de la G-6-Pasa es estimulada por PKA sin fosforilación. Por otro lado la glucógeno sintasa (GS) es fosforilada por PKA inhibiendo su actividad. Como resultado de todo lo anterior se estimula la glucogenólisis y se inhibe la glucogenogénesis. PKA fosforila a la fosfofructoquinasa 2 (PFK2), la cual en esta condición se inhibe su actividad de quinasa y se estimula su función de fructosa 2,6 bifosfatofosfatasa (FBPasa 2), trayendo como consecuencia una disminución de los niveles de fructosa 2,6 bifosfato (F2,6P2), metabolito que es un modulador alostérico negativo de la fructosa 1,6 bifosfatofosfatasa 1 (FBPasa 1) con lo cual se estimula la neglucogénesis. Por otro lado los bajos niveles de F2,6P2disminuye la actividad de la fosfofructoquinasa 1 (PFK1) trayendo como consecuencia inhibición de la glicolisis. La actividad de la fosfoenolpiruvato carboxiquinasa es estimulada por PKA lo cual incrementa la neoglucógenesis. Así mismo la PKA fosforila e inactiva a la piruvato quinasa hepática (PK), lo que contribuye a la disminución de la glicolisis.
|