Inmunología
Las Células de Langerhans. Los inmunocitos viajeros de la piel
Maduración y Migración
La mayoría de los estudios sobre la maduración de las células de Langerhans durante su migración hacia los ganglios linfáticos, se han realizado en cultivo debido a la dificultad de seguir el recorrido de estas células in vivo. Las células de Langerhans cultivadas por más de 72 horas, disminuyen su capacidad de fagocitar y procesar antígenos, pero adquieren una mayor capacidad para activar a linfocitos T. Estas observaciones sugieren que las células de Langerhans cultivadas o maduras son equivalentes a las células que en los ganglios linfáticos participan presentando antígenos en la fase de inmunoestimulación de la respuesta inmunitaria6. Este axioma ha permitido evaluar el proceso de migración y maduración de las células de Langerhans en cultivo.
Durante la maduración en cultivo, las CL aumentan su tamaño y la capacidad de estimular a las células T, mientras que su superficie sufre varios cambios. Aumenta la expresión de MHC clase II, B7, ICAM 1, LFA 3 y CD40. Otras moléculas como ICAM3, CD11b, CD45 también aumentan, mientras disminuye la expresión del receptor Fcg R y CD326.
Las células de Langerhans inmaduras (2-4 horas de cultivo) poseen una alta rata de biosíntesis de MHC-II, mientras que las células maduras (1-3 días de cultivo) manifiestan una bajoregulación de este proceso y el complejo MHC-péptido se hace mas estable en el tiempo, pudiéndose detectar a las 72 horas después de la exposición al antígeno. En el caso de antígenos de Leishmania major, este proceso solo ocurre cuando hay fagocitosis8. La disposición de las moléculas MHC clase II en las CL infectadas in vitro con Leishmania major, es diferente a lo largo del tiempo una ves que transcurre la infección. A las 8h las moléculas se encuentran en la vacuola parasitófora y en la periferia del núcleo, y a las 24 h solo se observan en la borde de la célula, sugiriendo que es entonces cuando la célula es capaz de presentar el antígeno.
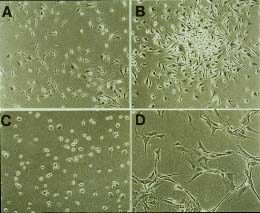
Línea de Células de Langerhans múridas inmadura
XS de Akira Takashima.
La maduración de las células de Langerhans es regulada por citocinas. Por ejemplo, el GM-CSF, producido por los queratinocitos activados, promueve la expresión de sus receptores en las células epidérmicas e induce a las células de Langerhans a madurar y migrar hacia los ganglios linfáticos regionales. Esta función la induce GM-CSF conjuntamente con TNF a , producida también por las células epidérmicas. Se ha visto que TNF a puede afectar la maduración disminuyendo la capacidad de procesar antígeno y la IL-1 aumentando la capacidad de presentar el mismo. La última actúa de forma paracrina y autocrina induciendo la expresión de sus receptores6.
La cinética de la migración in vivo de las células de Langerhans ha sido estudiada mediante la aplicación epicutánea del hapteno fluorescente Rodamina B en ratones9. Estos estudios demostraron un descenso rápido (aproximadamente 50%) en la densidad de células dendríticas epidérmicas, en las primeras 16 horas después de la sensibilización. La observación coincidió con la aparición de células dendríticas Rodamina B+ en el ganglio linfático adyacente. En el ganglio linfático, las primeras células portadoras del hapteno se observaron a las 6 horas y el mayor número entre 24-48 horas después de la sensibilización, representando un 6% del total de células dendríticas ganglionares.
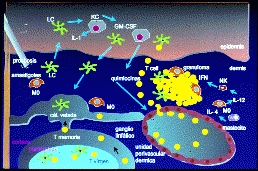
Respuesta inmunitaria cutánea frente a un antígeno:
Captura de antígenos, migración de células dendríticas
al ganglio linfático, sensibilización de linfocitos T vírgenes
, y establecimiento de la respuesta inflamatoria.
Los leucocitos utilizan ß2-integrinas para interactuar entre sí, y b 1-integrinas para interactuar con los componentes de la matriz extracelular10. Durante la travesía de la epidermis hacia los ganglios linfáticos, las células de Langerhans interactúan con laminina y colágeno IV en la membrana basal, con colágeno tipo I en la dermis superior, y con fibronectina en los linfáticos aferentes. La adhesión de las células de Langerhans a fibronecina y laminina es mediada por los receptores a5ß1 y a6ß1 respectivamente. Staquet y cols., (1995) demostraron que la interacción de las células de Langerhans con la matriz extracelular dérmica (colágeno tipo I) sólo se realiza después del contacto con componentes de la membrana basal (laminina, colágeno tipo IV)11. De igual manera, un contacto inicial de las células de Langerhans con la matriz dérmica reduce la capacidad de unión a la laminina de la membrana basal, interacción que impide el regreso de las células a la epidermis. Recientemente, Weiss y cols. (1997) demostraron que las células de Langerhans durante su migración expresan diferentes variantes genéticas de la molécula CD44. Esta molécula tiene gran importancia en la migración celular fisiológica o patológica al ganglio linfático12.
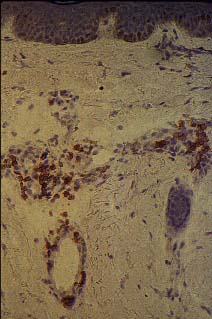 Extravasación y participación en proceso inflamatorio de leucocitos
positivos para el antígeno cútáneo linfocitario (CLA). |