Freddy González-Mujica
freddygonzalezmujica@gmail.com
Médico. Cirujano. PhD. en Bioquímica. Sección de Bioquímica Médica, Instituto de Medicina Experimental, Facultad de Medicina, Universidad Central de Venezuela. Caracas. Venezuela
Bioquímica Insulina. Estructura, síntesis, secreción, depuración y degradación (Revisión) Fecha de recepción: 19/06/2017
Fecha de aceptación:
28/09/2017
La insulina es una hormona polipeptídica formada por 2 cadenas, una de 21 aminoácidos, la A y otra de 30 aminoácidos, la B, unidas por 2 enlaces disulfuro y existe un tercer enlace disulfuro dentro de la cadena A. La estructura secundaria es muy compleja para el tamaño de la molécula, presentando estructura α helicoidal y giros β en ambas cadenas y lámina β en la cadena B. La forma activa de la insulina es monomérica, así está en la circulación general, es la que se une al receptor y existe a concentraciones de 10-6 M. A concentraciones mayores se forman dímeros por interacciones entre las cadena B. En los gránulos secretorios de las células β la insulina forma hexámeros coordinados con 2 átomos de Zn2+ y es la forma de almacenamiento de la hormona.
En humanos existe un solo gen de la insulina ubicado en el cromosoma 11p15.5 en el cual se encuentran 3 exones y 2 intrones. El ARNm maduro en 5 posee 7-metil guanosina y en 3 una cola de poliadenina, dicha molécula sirve de molde para la síntesis de la preproinsulina de 110 aminoácidos, ésta madura por la eliminación del péptido líder cuando entra al retículo endoplasmático dando origen a proinsulina la cual se pliega y forma los enlaces disulfuro antes de ser transferida al aparato de Golgi donde se elimina el péptido C dando la insulina la cual se almacena en gránulos secretorios. Existen controles transcripcionales y post-transcripcionales durante la síntesis de insulina y ambos procesos están bajo regulación.
La secreción de insulina puede depender de los canales de K+ATP, mecanismo mediante el cual se libera insulina y está en relación directa a la glicemia y al metabolismo de la glucosa. La producción de ATP corre paralelo con la utilización de la glucosa lo cual incrementa la relación ATP/ADP y esto condiciona el cierre de los canales de K+ATP despolarizándose la membrana plasmática con la apertura de los canales de Ca2+ incrementándose la concentración intracelular del mismo y secretándose la insulina. El mecanismo independiente de los canales de K+ATP es mediado por las incretinas (péptido insulinotropico dependiente de glucosa GIP y el peptido1 similar al glucagón GLP1), las cuales se unen a su receptor y mediante unas proteínas G incrementan la actividad de la adenilato ciclasa y en consecuencia se eleva la cantidad de AMPc y con ello la actividad de la proteína quinasa A y de Epac 2 liberándose la insulina. Los mecanismos de secreción de insulina están regulados de manera muy precisa y se adecuan a los requerimientos metabólicos del organismo.
La depuración de insulina ocurre en todas las células sensibles a la hormona pero en mayor medida en el hígado, riñón y músculo esquelético. La insulina unida al receptor es internalizada en vesículas endocíticas donde se inicia su degradación por la participación de una enzima específica que degrada insulina (IDE por sus siglas en inglés).
Palabras Claves:Insulina, estructura de insulina, síntesis de insulina, secreción de insulina, depuración de insulina, degradación de insulina, preproinsulina, proinsulina, incretinas, diabetes.
Title Insulin. Structure, Synthesis, Secretion, Clearence and Degradation
Abstract
Insulin is a polypeptide hormone formed by 2 chains, one of 21 amino acids, A and another of 30 amino acids, B, linked by 2 disulfide bonds and a third disulfide bond exists within the A chain. The secondary structure is very complex for the size of the molecule, presenting α helical structure and β-turns in both chains and β-sheet in the B-chain. The active form of insulin is monomeric, so it is in the general circulation, also it is the one that binds to the receptor and exists at concentrations of 10-6 M. At higher concentrations dimers are formed by interactions between the B chains. In the secretory granules of the β cell the insulin forms hexamers coordinated with 2 Zn2+ atoms and is the form of storage of the hormone.
In humans there is a single insulin gene located in the chromosome 11p15.5 in which 3 exons and 2 introns are found. Mature mRNA in 5' possesses 7-methyl guanosine and in 3' a polyadenine tail, which serves as a template for the synthesis of 110 amino acids preproinsulin. IT mature by eliminating the leader peptide when it enters the endoplasmic reticulum giving rise to proinsulin which folds and forms the disulfide bonds before being transferred to the Golgi apparatus where the C-peptide is removed giving the insulin which is stored in secretory granules. There are transcriptional and post-transcriptional controls during insulin synthesis and both processes are under regulation.
Insulin secretion may depend on K+ATP channels, a mechanism by which insulin is released in direct relation to glycaemia and glucose metabolism. The production of ATP is parallel to glucose utilization, which increases the ATP / ADP ratio and this condition closure of the K+ATP channels depolarizing the plasma membrane with the opening of the Ca2+ channels, increasing its intracellular concentration and secreting the insulin. The mechanism independent of K+ATP channels is mediated by incretin (glucose-dependent insulinotropic peptide GIP and glucagon-like peptide1GLP1-), which bind to its receptor and by G proteins increase the activity of adenylate cyclase and consequently increases the amount of cAMP and thus the activity of protein kinase A and of Epac 2 are increased, releasing the insulin. The mechanisms of insulin secretion are regulated very precisely and are adapted to the metabolic requirements of the body.
The clearance of insulin occurred in all cells sensitive to the hormone, but it is higher in liver, kidney and skeletal muscle. Insulin bounds to its receptor is internalized in endocytic vesicles where its degradation start by the action of a specific enzyme that degraded insulin (IDE).
Insulina. Estructura, síntesis, secreción, depuración y degradación (Revisión)
Introducción
La insulina es una hormona
polipeptídica anabólica de 51 aminoácidos, secretada por las células β de los
islotes de Langerhans, la cual consta de dos cadenas polipeptídicas
designadas A y B conectadas por enlaces
disulfuro. Una de las funciones primarias de la hormona es la de incrementar la toma de glucosa desde la sangre por los tejidos
muscular y adiposo, y la reducción de la producción hepática de glucosa lo cual
en su conjunto se traduce en la regulación de la homeostasis de la glucosa y en
la prevención de la diabetes mellitus (1). La diabetes se
caracteriza por una disminución de la tolerancia a la glucosa como resultado de
una deficiencia relativa de la producción de insulina o una falta de
sensibilidad a la hormona o a una combinación de ambas, lo cual trae como
consecuencia hiperglicemia y esta última generalmente se asocia con
complicaciones tales como enfermedad vascular, en particular coronariopatía,
enfermedad vascular cerebral, retinopatía, nefropatía y neuropatía (2).
En 1890 se demostró que la
pancreatectomía condiciona el desarrollo de diabetes (3). Schafer en
1916 (4), especuló sobre la
existencia de una hormona antidiabética producida por los islotes pancreáticos
la cual designó como insulina; poco después se
demostró que la ligadura del conducto pancreático conduce a la destrucción del
páncreas exocrino y solo se produce diabetes si los islotes de Langerhans son
destruidos también(5). Posteriormente los trabajos de Banting, Best,
Collip y MacCleod(6) condujeron al descubrimiento de la insulina la
cual no solo controla la homeostasis de la glucosa sino también participa en la
regulación del metabolismo intermediario de lípidos y proteínas, en la síntesis
de ARN y ADN y en el crecimiento y diferenciación celular.
En el presente trabajo
pasaremos revista a las características estructurales de la insulina, al
mecanismo de su síntesis y maduración, a los aspectos relacionados con su
secreción, depuración y degradación.
Estructura de la insulina
La insulina monomérica
consta de dos cadenas polipeptídicas: la cadena A está formada por 21
aminoácidos y la cadena B por 30 aminoácidos unidas por enlaces disulfuro y con
un peso molecular de 5800 Daltons. El monómero posee 3 enlaces disulfuro, dos
de ellos entre las cadenas A y B (A7 con B7 y A20 con B19) y uno dentro de la
cadena A (A 6 con A11) (7). Sanger
(8)y su grupo
establecieron la secuencia de aminoácidos de ambas cadenas polipeptídicas así
como la ubicación de los enlaces disulfuro, lo cual se muestra en la Figura 1.
Figura
1. Secuencia
de aminoácidos de la insulina humana.Se
esquematiza en azul la secuencia de aminoácidos de la cadena A (21 aminoácidos)
y en violeta los de la cadena B (30 aminoácidos) de la insulina humana. Se
destacan en naranja los grupos amino y carboxilo terminales de los aminoácidos
iniciales y finales de cada cadena, también se destacan en naranja los enlaces
disulfuro entre las cadenas A y B (A7 con B7 y A20 con B19) y entre la cadena A
(A6 con A11). Para detalles ver el texto.
La estructura secundaria de
la Cadena A consta de 2 α hélices dispuestas antiparalelamente, la primera formada
entre los aminoácidos A2 al A8 y la segunda entre los aminoácidos A13 al A19,
las dos hélices están conectadas por el segmento de aminoácidos A9 Al A12, el
cual está dispuesto en un giro en U de tal suerte que ambas hélices se
encuentran lado a lado y el extremo amino terminal de una de ellas está próximo
al extremo carboxilo terminal de la otra (ver Figura 2) (9).
La estructura secundaria
de la cadena B presenta α hélice y lámina β y cuando se cristaliza existe en
dos configuraciones (10). En la configuración T existe una α hélice
central entre los aminoácidos B9 al B19, la cual muestra un patrón de enlaces
de hidrógeno del aminoácido 1 al 5 en lugar del patrón clásico 1 al 4. A
continuación de la hélice se encuentra un giro β entre las Gly B20 y B23
permitiendo un plegado en forma de U. Los residuos de los aminoácidos B24 al
B30 forman una lámina β extendida, la cual gracias al giro β se aproxima a la α
hélice central con la Phe B24 y la Tyr B26, de la lámina β, en intimo contacto
con las Leu B11 y B15, de la α hélice central (Figura 2). En la configuración R
existe una α hélice desde B1 hasta B19 con el resto de la estructura como se
describió antes (9). Los enlaces disulfuro entre las Cys A7 y B7 y
A20 y B19 contribuyen a estabilizar la estructura nativa de la insulina. La
estructura secundaria de ambas cadenas A y B es sorprendentemente compleja para
un péptido pequeño y esas intrincadas interacciones entre las cadenas laterales
de los aminoácidos contribuyen a determinar la afinidad de la hormona por su
receptor.
Figura
2. Estructura
secundaria de la insulina.Se
esquematiza en verde la estructura secundaria de la cadena A destacándose las
dos α hélices antiparalelas como cintas y el giro β que las une. La cadena B
está esquematizada en azul, la porción de α hélice central se destaca como una
cinta y la estructura en lámina β como una flecha en forma de cinta. Se destacan
en naranja los enlaces disulfuro,
también se destacan los grupos amino y carboxilo terminales de ambas cadenas.
El esquema corresponde a la configuración T.
La estructura monomérica de
la insulina existe a concentraciones de aproximadamente 10-6 M, es
la forma que está presente en la circulación general y es la forma activa de la
hormona que se une a su receptor. A concentraciones superiores (≈ 10-5 M),
como la que se encuentra en la circulación portal, la insulina forma dímeros,
gracias al establecimiento de puentes de hidrógeno y enlaces hidrofóbicos entre
los aminoácidos de los extremos carboxilo terminales de las cadenas B, en
configuración de lámina β, de dos moléculas de insulina (11).
La insulina se deposita en
las células β de los islotes pancreáticos en gránulos densos, en los cuales se
encuentra la insulina en forma cristalina insoluble y hexamérica, la
concentración de insulina en dichos gránulos es de aproximadamente 40 mM (11).
En el hexámero las 6 moléculas de insulina se agrupan en 3 dímeros, además se
coordinan con 2 átomos de Zn2+ a través del grupo imidazol de 3
histidinas (His B10) y también el Zn2+ forma enlaces de coordinación
con 3 moléculas de agua. En el hexámero las 6 moléculas de insulina están en la
configuración T antes descrita (11) (Figura 3). Por lo antes mencionado
se puede afirmar que la configuración monomérica de la insulina es la forma
activa de la hormona y que el hexámero es la configuración de depósito.
Figura 3. Configuración hexamérica
de la insulina. En el esquema se
muestran los 6 monómeros de insulina con diferentes colores agrupados en 3
dímeros, cada uno de los cuales forma un enlace de coordinación, a través de la
His B10, con los 2 átomos de Zn2+ centrales
En la insulina se describen
2 sitios de unión al receptor, el sitio 1 o superficie de unión clásica
incluye aminoácidos de ambas cadenas, siendo los de la cadena A: A1 Gly, A5
Gln, A19 Tyr y A21 Asn y los residuos de la cadena B son: B12 Val, B16 Tyr B24,
Phe, B25 Phe y B26 Tyr. En sitio 2 están incluidos también aminoácidos de ambas
cadenas: Ser A12, Leu A13, Glu A17, His B10, Glu B13 y Glu B17. En
general se acepta que el sitio 1 de la insulina se une al sitio 1 de receptor
de la hormona y el sitio 2 de la insulina se une al sitio 2 del receptor (11).
Biosíntesis de la insulina.
La insulina es una hormona
polipeptídica de 51 aminoácidos, sin embargo se sintetiza como un precursor de
110 aminoácidos denominado preproinsulina, cuya existencia fue demostrada por Chang
y colaboradores(12), en el mismo están unidas sucesivamente las
secuencias de aminoácidos de un péptido líder, la cadena B, el péptido C y la
cadena A (ver más adelante. Figura 5)
En los humanos existe una copia del gen de la insulina, Harper
y colaboradores (13) demostraron mediante el uso de hibridación in situ, que el gen que codifica para la
insulina está ubicado en el brazo corto del cromosoma 11 (11p15.5). Casi
simultáneamente se reportó la secuencia de dicho gen(14) estableciéndose
que el mismo está constituido por 3 exones (región codificante de un gen o
preARNm) y 2 intrones (región no codificante de un gen o preARNm). El exón 1
codifica para una región del ARNm (ácido ribonucleico mensajero) que no se
traduce, el exón 2 codifica al péptido señal a la cadena B y parte del péptido
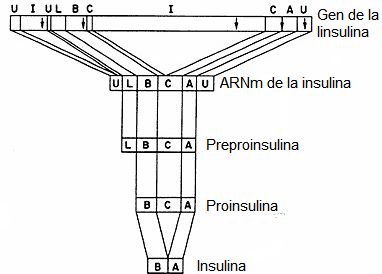
C y el exón 3 codifica al resto del péptido C y la cadena A. (Figura 4).
Figura
4. Expresión
del gen de la insulina.Se
esquematizan de arriba abajo la estructura del gen de la insulina, el ARNm
maduro, la proteína traducida a partir del mismo, preproinsulina y su
maduración hasta producir insulina. Las letras corresponden a: I a los
intrones, U a las regiones no traducidas, L al péptido líder, B a la cadena B,
C al péptido C y A a la cadena A.
La primera etapa en la
síntesis de la insulina ocurre a nivel de los ácidos nucleicos, el ARN
transcrito primario mediante el mecanismo del splicing (eliminación de
intrones y empalme de exones) pierde los dos intrones, en el extremo 5 se le
adiciona 7-metil guanosina (casquete) y en el extremo 3 se le añade la cola de
poliadenina (poli A) obteniéndose el ARMm maduro el cual es traducido en el
retículo edoplamático rugoso para dar preproinsulina.
Como ocurre con otras
proteínas secretadas, la preproinsulina (Figura 5) contiene un péptido señal o
líder amino terminal hidrofóbico de 24 aminoácidos, el cual al ser sintetizado
en los ribosomas y antes de concluir la síntesis total de la proteína, se une
en el citosol a partículas ribonucleoproteicas que reconocen al péptido señal
(SRP por sus siglas en inglés) (15). Dichas partículas facilitan que
la preproinsulina sea translocada a través de la membrana del retículo
endoplamático hasta la cisterna del mismo. Este proceso ocurre por medio de un
canal especializado que conduce péptidos (16), una vez en la
cisterna del retículo endoplasmático, el péptido señal de la preproinsulina es
hidrolizado gracias a la participación de una peptidasa señal generándose la
proinsulina (17).
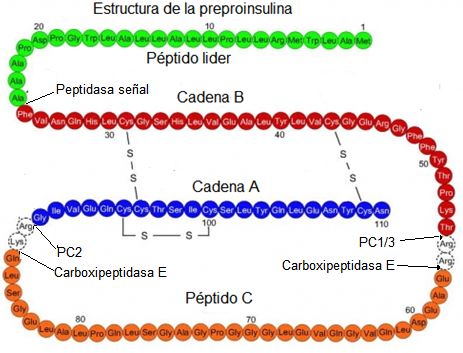
Figura
5. Estructura
de la preproinsulina humana.En el
esquema se representa la secuencia de aminoácidos de la preproinsulina humana.
En verde se destaca el péptido líder o señal, en rojo la cadena B, en naranja
el péptido C, en azul la cadena A y en transparente los aminoácidos de ambos
extremos del péptido C que son eliminados durante el procesamiento de la
proinsulina. Se destacan los sitios de acción de las proteasas: peptidasa
señal, PC2, PC1/3 y carboxipeptidasa E
El péptido señal es prontamente
degradado y en consecuencia no es un producto de secreción de las células
β. A continuación la proinsulina se
pliega y ocurre la formación de los tres enlaces disulfuro (18)
proceso que requiere una variada gama de proteínas chaperonas del retículo
endoplasmático tales como la reductasa de grupos tioles de las proteínas (19).
Luego de adquirir el plegamiento tridimensional, la proinsulina es transferida
del retículo endopasmático al aparato de Golgi y entra a vesículas secretoras
inmaduras.
La proinsulina a pesar de su
mayor tamaño comparte varias de las características físicas de la insulina, la
proinsulina forma dímeros y hexámeros de coordinación con el Zn2+,
tienen puntos isoeléctricos similares, igual solubilidad y reacción cruzada con
los anticuerpos. Estos hallazgos sugieren que la estructura de la insulina en
la proinsulina esta conservada y que el péptido C, de unos 35 aminoácidos,
tiene una configuración flexible pero no completamente desordenada.
La conversión de proinsulina
en insulina se inicia en el trans Golgi y se acelera en los gránulos
prosecretarios en la medida en que estos
se acidifican y maduran, la misma tiene lugar por la participación de unas
endopeptidasas similar a la tripsina y
una exopeptidasa similar a la carboxipeptidasa B. En los gránulos secretorios
existen dos endoproteasas que participan en la conversión de proinsulina en
insulina denominadas PC2 y PC1/3, la primera hidroliza después de los
aminoácidos básicos Lys-Arg en la unión péptido C/cadena A y la segunda
preferentemente hidroliza antes del dipéptido Arg-Arg en la unión cadena B/
péptido C aun cuando puede actuar en ambos extremos (Figura 5). Ambas
endopeptidasas presentan un pH óptimo cercano a 5,5 consistente con el
encontrado en los gránulos secretores de las células β, el cual se alcanza
gracias a la participación de una bomba de protones presente en dichos gránulos
secretorios (20). La enzima que elimina los dipéptidos básico
después de la acción de PC2 y PC1/3 se denomina carboxipeptidasa E (Figura 5).
Los estudios cinéticos han demostrado que primero actúa PC1/3 generando un
intermediario y luego lo hace PC2 y por último la carboxipeptidasa E. A
diferencia del péptido líder, el péptido C no es hidrolizado y es secretado
junto con la insulina en cantidades equimolares.
La insulina recién
sintetizada forma cristales con el Zn2+, el cual es transportado al
interior de los gránulos por un transportador denominado ZnT8 (21),
dichos cristales se acumulan en el centro electrón denso de los gránulos y en
la periferia menos densa se encuentra el péptido C.
Las insulinopatias clásicas:
insulina Chicago en la cual se sustituye la Leu B25 por Phe (22) ;
insulina Los Ángeles en la cual se reemplaza la Phe B24 por Ser (23)
e insulina Wakayama en la cual se sustituye Val A3 por Leu (23)
resultan de mutaciones puntuales acumulándose
en la sangre las insulinas mutantes las cuales presentan una menor capacidad
para unirse al receptor de insulina y en consecuencia una menor actividad
biológica. Síndromes clínicos raros resultan de mutaciones en otros sitios que
afectan etapas claves en la biosíntesis de la insulina (23).
Regulación de la síntesis de insulina
La biosíntesis de la
insulina está regulada tanto a nivel de la transcripción como de la traducción.
Las secuencias de señales que determinan la exclusividad de la expresión del
gen de la insulina en las células β de los islotes pancreáticos están ubicadas
entre - 520 y + 1 pares de bases relativas al inicio de la transcripción (24).
En las células β de los islotes pancreáticos
existen unos 13.000 gránulos secretorios los cuales ocupan aproximadamente un
10 % del volumen celular y cada gránulo contiene cerca de 200.000 moléculas de
insulina (25). Sin embargo el contenido de insulina de las células β
es altamente dinámico, acumulándose la hormona en la presencia de nutrientes y
disminuyéndose en la ausencia de los mismos. La habilidad de las células β de
responder rápidamente a señales celulares, generalmente está relacionado con
regulación post-transcripcional. En la región promotora del gen le la insulina
existen un número de secuencias de bases denominadas elementos A, C, E, Z y el
elemento que responde al AMPc (CRE por sus siglas en inglés) que determinan la
localización de la insulina en las células β, así como también la unión a
diferentes factores de transcripción los cuales determinan la regulación de la
expresión genética de la insulina(26). La región promotora del gen
de la insulina se extiende aproximadamente a unas 400 pares de bases antes del
punto de inicio de la transcripción(26).
Es interesante mencionar que
uno de los factores de transcripción que une al elemento C es denominado MafA
el cual se expresa exclusivamente en las células β, con lo cual condiciona, al
menos en parte, la exclusividad de la expresión del gen de la insulina en
dichas células(27) y además media la regulación de la expresión del
gen de la insulina por la glucosa.
En respuesta a la presencia
de nutrientes, las células β incrementan la síntesis proteica, por lo menos en parte, por la
desfosforilación del factor eucariota de iniciación 2a (eIF2a por sus siglas en
inglés) mediante la participación de la proteína fosfatasa 1 la cual es estimulada
por glucosa (28), por el contrario la quinasa pancreática del
retículo endoplasmático fosforila a eIF2a regulando negativamente la
traducción.
En las células β existe un
mecanismo que permite detectar la cantidad de insulina almacenada y secretada y
consecuentemente ajustar su síntesis. La proteína granular transmembranosa
denominada autoantígeno de las células de los islotes 512 (ICA512 por sus
siglas en inglés) es una parte esencial del mecanismo de control por retroalimentación.
Los gránulos de insulina se fusionan transitoriamente con la membrana
plasmática para liberar insulina y simultáneamente la elevada concentración de
Ca2+ activa una proteasa µ calpain la cual hidroliza el segmento
citoplamático de ICA512; éste último migra al núcleo donde se une al factor de
transcripción STAT5, impidiendo su defosforilación y regulando positivamente la
expresión del gen de la insulina (29). En consecuencia la liberación
de insulina contenida en los gránulos secretores es comunicada al núcleo donde
funciona como un mecanismo de retroalimentación positiva iniciándose la
síntesis de insulina con lo cual se mantiene una cantidad adecuada de la
hormona en depósito.
La velocidad de la
traducción del ARNm de la preproinsulina se incrementa cuando los islotes
pancreáticos son incubados con altas concentraciones de glucosa (25 mM) evento
que es independiente de la transcripción (30).
Los resultados de los
estudios in vitro indican que la
estabilidad del ARNm de la preproinsulina disminuye en condiciones de una baja
concentración de glucosa y por el contrario se incrementa con la elevación de
la concentración de la hexosa (31).
Las proteínas que unen
polipirimidinas, regiones ricas en uridina y citosina en la región 5 no traducida
del ARNm, incrementan la viabilidad del ARN y estimulan el inicio de la
traducción no solo de la preproinsulina sino también de otras proteínas de los
gránulos secretorios tales como la ICA512 y la PC2. Es interesante mencionar
que en la región 5 no transcrita del ARNm de la preproinsulina existe una
secuencia de bases que juega un papel esencial en la regulación de la
traducción, ya que su remoción bloque la estimulación de la síntesis de
preproinsulina por la glucosa (32).
La regulación
post-transcripcional modula la síntesis de la insulina de manera inmediata, por
el contrario la regulación transcripcional modifica la síntesis de insulina a largo
plazo.
Secreción de insulina
En
los sujetos sanos la liberación de la insulina está exactamente controlada para
alcanzar las demandas metabólicas, las células β detectan los cambios en la
glicemia y liberan la cantidad exacta de insulina (33). Para
detectar el estado nutricional las células β están agrupadas en islotes los cuales
están conectados estratégicamente con los vasos sanguíneos. Los islotes forman
una densa red con los vasos sanguíneos pequeños y reciben 10 veces más sangre
que el tejido exocrino circundante. Los capilares que irrigan los islotes están
fenestrados, estructura que incrementa la permeabilidad capilar, lo cual
permite un íntimo contacto de las células β con los nutrientes presentes en la
sangre, así mismo facilita que la insulina secretada alcance la circulación (34).
Además de la glucosa, algunos aminoácidos y ácidos grasos pueden regular la
secreción de insulina.
La secreción de insulina se
realiza mediantes dos mecanismos: uno relacionado con los canales de K+
dependientes de ATP y otro que es independiente de dichos canales. A
continuación discutiremos ambos mecanismos.
Secreción de insulina dependiente de canales de K+ATP.
Los
islotes de Langerhans son pequeños órganos encargados de detectar los cambios
en las cantidades de los nutrientes y hormonas presentes en el medio ambiente
que los rodea, además de responder a estímulos nerviosos. El aparato secretor
de insulina de las células β está equipado con controles metabólicos en
diferentes etapas de señalización que están bajo riguroso control. La
maquinaria metabólica de las células β está diseñada para detectar las
variaciones de la glicemia y liberar insulina de acuerdo a los requerimientos
el organismo(35). Además de la glucosa, algunos aminoácidos
incluyendo glutamina y leucina, así como los ácidos grasos son capaces de
estimular la secreción de insulina en respuesta a la glucosa (35,36).
La estimulación de la secreción de insulina en la fase temprana pre-absortiva
es mediada por la inervación parasimpática de los islotes (37.
El
metabolismo de la glucosa en las células β, corre paralelo con el incremento en
la producción de ATP y el consecuente aumento de la relación ATP/ADP lo cual
condiciona el cierre y la inhibición de los canales de potasio dependientes de
ATP (K+ATP) despolarizándose la membrana plasmática. Los canales de K+ATP
son un complejo constituido por 4 subunidades del receptor sensibles a las
sulfonilureas 1 (SUR 1, por sus siglas en
inglés), las cuales son las subunidades regulatorias sensibles a ATP y que se
encuentran rodeando a 4 subunidades del canal iónico de potasio (Kir6.2)
propiamente dicho. Cuando la relación ATP/ADP se incremente la subunidad SUR1
une ATP cerrando el canal iónico de K+ con lo cual se incrementa la
concentración intracelular del catión, despolarizándose la membrana plasmática.
En respuesta a la despolarización de la membrana plasmática por el cierre de
los canales de K+ATP se
abren los canales de Ca++ tipo L dependientes de voltaje y se
produce un influjo de Ca++ lo
cual es conocido como uno de los eventos primarios en la exocitosis de la
insulina (Ver Figura 6) (38).
La
habilidad de las células β de responder a las fluctuaciones de la glicemia en
un rango comprendido ente 3 y 16 mM se puede realizar gracias al concurso
de dos proteínas; la primera de ellas es el transportador de glucosa
independiente de Na+ (GLUT 1 en el hombre y GLUT 2 en roedores) que
presenta un alto KM para la glucosa (≈ 17 mM) lo cual permite un
rápido equilibrio de la concentración de glucosa intra y extra celular; la otra
es la hexoquinasa IV o glucoquinasa, la cual cataliza la primera reacción de la
utilización de la glucosa y en particular de la glicólisis con un KM para
la glucosa de ≈ 10 mM(39). La combinación de la participación del
GLUT 1 y de la glucoquinasa condicionan un incremento de la glicólisis y del ATP, casi paralelamente con
el incremento de la glicemia y en consecuencia una liberación de insulina
proporcional al cambio en la concentración de glucosa en sangre (35).
En las células β la glicolisis y el ciclo de Krebs están estrechamente
relacionados por la baja expresión genética de la lactato deshidrogenasa lo
cual permite aún más un paralelismo entre la glicemia, la producción de ATP y
la secreción de insulina (40).
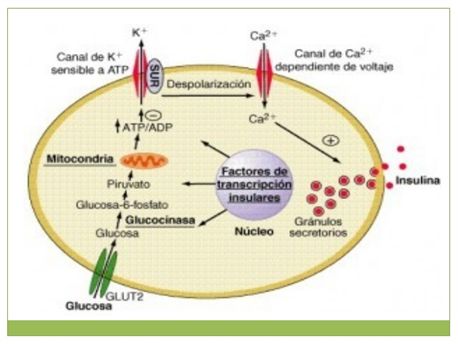
Figura 6. Mecanismo
de la secreción de insulina por las células β pancreáticas dependiente de los
canales K+ATP. La utilización de la glucosa, por las
células β corre paralela con la glicemia gracias a la participación del GLUT 2
(en roedores) y de la glucoquinasa, con lo cual la síntesis de ATP está
relacionada con la glicemia. El incremento de la relación ATP/ADP cierra los
canales de K+ATP despolarizándose la membrana plasmática lo
cual condiciona la apertura de los canales de Ca++ produciéndose la
liberación de insulina. Para detalles ver el texto.
La
secreción de insulina está orquestada por varios factores, evidentemente el Ca++
es uno de ellos, además existen efectores proteicos de la exocitosis
asociados a las vesículas (un factor soluble sensible a
N-etilmaleimida que se une a un receptor proteico, SNARE por sus siglas en
inglés), que contienen insulina, y a
la membrana plasmática lo cual facilita la fusión entre ambas (41).
La secreción de insulina transcurre en dos fases,
la primera consiste de un pico inicial que ocurre entre 3 y 10 minutos de la
ingesta de alimentos y una segunda fase de desarrollo más lento; la primera
fase está disminuida en los pre-diabéticos y está casi totalmente ausente en los
diabéticos tipo 2 con una disminución
variable de la segunda fase (42). De los aproximadamente 13.000
gránulos de insulina que existen en la
célula β unos 500 están adosados a la membrana plasmática y
de estos unos 100 muy próximos a los canales de Ca++ y que son los
que contribuyen a la primera fase de secreción; una vez que estos han liberado
la insulina, son reemplazados por el reclutamiento de otros gránulos
produciéndose la segunda fase más sostenida (43).
Secreción
de insulina independiente de los canales de K+ATP
Las
incretinas son hormonas producidas en el intestino en respuesta a la ingesta de
alimentos, y que han sido reconocidas como estimuladoras fisiológicas de la
secreción de insulina(44). El polipéptido insulinotrópico dependiente de glucosa (GIP por sus siglas en
inglés) es secretado por las células K, ubicadas en la parte proximal del
intestino delgado y el péptido 1 similar al glucagón (GLP 1 por sus siglas en
inglés) es producido por las células L ubicadas en la porción distal del
intestino delgado y el colon (45).
Tanto
GIP como GLP 1 se unen, en las células β pancreáticas, a un receptor de
membrana constituido por 3 subunidades, el cual por medio de una proteína G
estimula la adenilato ciclasa, ésta enzima a su vez incrementa la concentración
intracelular de AMPc y éste estimula la secreción de insulina por un mecanismo dependiente de la vía de la proteinquinasa A (PKA por sus
siglas en inglés) y otro dependiente de una proteína intercambiadora estimulada
directamente por AMPc 2 (Epac 2 por sus siglas en inglés). El mecanismo por el cual la vía de PKA estimula la secreción de
insulina no está claro pero es independiente del cierre de los canales de K+ATP
y la subsecuente despolarización de las células β y el incremento de Ca++
intracelular, pero si está estrechamente relacionado con la concentración
sanguínea de glucosa (Ver Figura 7) (46).
Por otro lado, Epac 2 se une a AMPc y funciona como un factor intercambiador de
nucleótidos de guanina para las proteínas de bajo peso molecular similar a Ras,
denominada Rap 1. La interacción de Epac 2 con Rap 1 es un evento crítico para
promover la exocitosis de las vesículas que contienen insulina; además Epac 2
interactúa con una proteína de andamiaje denominada Rim 2 la cual está
localizada tanto en la membrana de las vesículas secretoras como en la membrana
plasmática, lo cual permite las etapas de acoplamiento e iniciación de la
exocitosis (47).
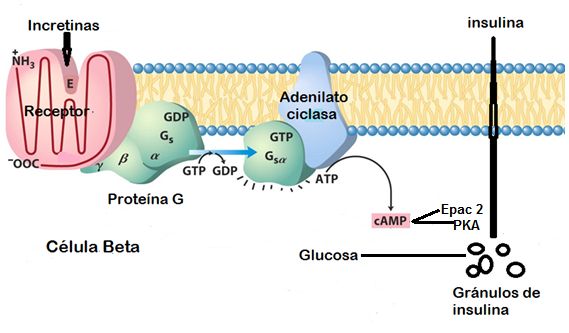
Figura 7. Secreción
de insulina por las células β mediada por incretinas e independiente de los
canales K+ATP. Las
incretinas se unen a receptores de membrana en las células β los cuales por
modulación de proteínas G condicionan el incremento en AMPc y éste estimula la
proteína quinasa A y a Epac 2, aumentándose la liberación de insulina por un
mecanismo desconocido pero dependiente de glucosa.
Regulación de la secreción de insulina
a.-
Glucosa. Éste carbohidrato es el estímulo primario para la liberación de
insulina en muchos animales incluyendo el hombre; en humanos, la ingesta de 75
g de glucosa incrementa los niveles de insulina desde el basal, de 20-30
pmol/L, hasta 250-300 pmol/L en 30 minutos, mientras que una ingesta similar de
lípidos o de lípidos más proteínas solo incrementa los valores de insulina hasta
50-60 pmol/L (48).
Las células β no cuentan con
un receptor de membrana para la glucosa que les permita detectar los cambios de
concentración; sin embargo el mecanismo antes descrito (Secreción de insulina
dependiente de los canales de K+ATP) les permite a las
células β adecuar finamente la secreción de insulina a la glicemia.
La ingesta de una cantidad
de glucosa incrementa más la secreción de insulina que la administración
intravenosa de una cantidad similar de glucosa (49) debido a la
liberación de las incretinas por las células del intestino que incrementa la
secreción de insulina (Secreción de insulina independiente de los canales de K+ATP).
b.- Aminoácidos. En general
los aminoácidos individuales son pobres secretagogos de insulina, sin embargo
algunas combinaciones de aminoácidos, a concentraciones fisiológicas o
superiores pueden estimular la secreción de insulina tal es el caso de la combinación
de glutamina y leucina (50).
c.- Ácidos grasos.
Recientemente se ha demostrado que las células β tienen receptores para los
ácidos grasos y que por medio de los mismos dichas moléculas influencian la
liberación de insulina (51).
d.- Estrógenos. Las células
β no son blancos clásicos de los estrógenos, sin embargo en dichas células se
encuentran receptores estrogénicos. La principal consecuencia fisiológica de la
acción del 17 β estradiol es el incremento de la secreción de insulina (52).
e.- Melanotonina. Es la
hormona producida por la glándula pineal, la misma atenúa la liberación de
insulina por las células β probablemente por disminución de la producción de
AMPc (53).
f.- La leptina es secretada por el tejido adiposo
y es conocida la influencia que tiene sobre el efecto de la insulina en los
tejidos adiposo y hepático. Generalmente se acepta que tiene un efecto
inhibitorio sobre la liberación de insulina (54).
g.- Hormona de crecimiento.
Una de las acciones mejor conocidas de la hormona de crecimiento es la
estimulación de la síntesis del factor I de crecimiento similar a la insulina,
el cual disminuye los niveles séricos de insulina y péptido C en humanos (55).
H.- Acetilcolina y
colecistoquinina. Este neurotransmisor y la hormona producida por el duodeno
potencian la secreción de insulina mediante el catabolismo del fosfoinositol (9).
Ambas moléculas se unen a sus respectivos receptores en la membrana plasmática
de las células β y mediante proteínas G activan la fosfolipasa C, la cual hidroliza
al fosfatidilinositol 4,5-bisfosfato (PPI2 por sus siglas en inglés)
produciendo inositol 1,4,5-trifosfatto (IP3 por sus siglas en inglés) y
diacilglicero, los cuales actúan como segundos mensajeros liberando calcio del
retículo endoplasmático(9). El incremento del calcio citosólico incrementa la liberación de insulina como se
describió antes (Secreción de insulina dependiente de los canales de K+ATP).
Depuración
y degradación de insulina
La captura y degradación de
la insulina es una característica de todos los tejidos sensibles a la hormona
(56). A concentraciones fisiológicas, la captura de insulina está mediada
por el receptor con una mínima participación de procesos no específicos. La
vida media de la insulina es de 4 a 6 minutos, como se pudiera esperar de una
rápida respuesta a los cambios de la
glicemia (56).
El hígado es el principal
órgano depurador de insulina, es capaz de captar el 50% de la hormona presente
en la circulación portal (57). La depuración hepática de la insulina
está disminuida en la diabetes y la obesidad (58). El riñón depura
el 50% de la insulina de la circulación general y el 70% del péptido C
circulante por filtración glomerular, reabsorción y degradación (59).
Además de hígado y riñón el tejido muscular juega un papel importante en la
depuración de insulina (58).
En condiciones normales la
casi totalidad de la insulina es degradada intracelularmente o por lo menos en
procesos que ocurren a nivel de la membrana plasmática (58). La
etapa inicial, en la toma de la insulina por las células, es la unión de la
hormona a su receptor, constituyendo un reservorio de insulina la cual puede
regresar a la circulación o ser internalizada (60). La insulina
unida al receptor es internalizada en vesículas endocíticas, donde puede
ocurrir el inicio de la degradación de la hormona gracias a la participación de
una enzima específica que degrada insulina (IDE por sus siglas en inglés) o ser
transferida intacta a otros organelos intracelulares como el núcleo, el aparato
de Golgi, el citosol, entre otros (58) o su liberación de la célula
intacta por diacitosis o retroendocitosis 58). La degradación de la
insulina se puede considerar como un mecanismo de terminar su acción.
Referencias
1.Bliss M. 1982. The Discovery of Insulin. Chicago:
University of Chicago Press 6.
2.Leslie RGD and Robbins DC 1995 Diabetes: Clinical
Science in Practice, Cambridge University Press 221-271.
3.Von Mering J and Minkowski O 1890 Diabetes mellitus
nach pankreas exterpation. Arch Exp Path Pharmacol (Leipzig) 26:371.
4.Schafer EA 1916. An introduction to the study of
internal secretion. In: The Endocrine Organs. London: Longmans, Gree.
5.Barron M. 1920. The relation of the islets of
Langerhans to diabetes with special referenceto cases of pancreatic lithiasis.
Surg Gynecol Obstet 31:437-448.
6.Banting FG and Best CH. 1921. The internal
secretion of the pancreas. J Lab Clin Med 7:247-253
8.Sanger F. 1959. Chemistry of insulin; determination of
the structure of insulin opens the way to greater understanding of life
processes.Science. 129: 1340-1344
9.Fu Z. Gilbert ER. and Liu D. 2013. Regulation of
Insulin Synthesis and Secretion and Pancreatic Beta-Cell Dysfunction in
Diabetes. Curr Diabetes Rev. 9: 2553.
10.Baker EN, Blundell TL, Cutfield JF, Cutfield SM,
Dodson EJ, Dodson GG, Hodgkin DC, Hubbard RE, Isaacs NW, Reynolds CD, Sakabe K,
Sakabe N, Vijayan M. 1988. The structure of 2 Zn pig insulin crystals at 1.5 A
resolution. Philos Trans R Soc London. B319:369456.
11.De Meyts P. 2004.Insulin and its receptor: structure,
function and evolution. Bioessays. 26: 135162.
12.Chan SJ, Keim P, and Steiner DF. 1976. Cell-free
synthesis of rat preproinsulins: characterization and partial amino acid
sequence determination. Proc. Natl. Acad. Sci. U. S. A. 73: 1964-1968.
13.Harper ME. Ullrich A. and Saunders GF. 1981.
Localization of the human insulin gene to the distal end of the short arm of
chromosome 11. Proc. Nat. Acad. Sci. 78: 4458-4460,
14.Bell, G.I., Pictet, R.L., Rutter, W.J., Cordell, B.,
Tischer, E., and Goodman, H.M. 1980. Sequence of the human insulin gene. Nature
284:26-32.
15.Egea PF, Stroud RM and Walter P. 2005.Targeting
proteins to membranes: structure of the signal recognition particle. Curr Opin
Struct Biol. 15: 21320.
16.Lomedico PT, Chan SJ, Steiner DF and Saunders GF.
1977. Immunological and chemical characterization of bovine preproinsulin. J
Biol Chem. 252: 79717978.
17.Patzelt C, Labrecque AD, Duguid JR, Carroll RJ, Keim
PS, Heinrikson RL, and Steiner DF. 1978. Detection and kinetic behavior of
preproinsulin in pancreatic islets. Proc Natl Acad Sci U S A. 75: 12601264.
18.Huang XF and Arvan P. 1995. Intracellular transport of
proinsulin in pancreatic beta-cells. Structural maturation probed by disulfide
accessibility. J Biol Chem. 270: 2041720423.
19.Munro S and Pelham HR. 1987. A C-terminal signal
prevents secretion of luminal ER proteins. Cell. 48: 899907.
20.Smeekens SP, Albiges-Rizo C, Carroll R, Martin S,
Ohagi S, Phillips LA, Benig M, Gardner P, Montag AG, Swift HH, Thomas G and
Steiner DF 1992 Proinsulin processing by the subtilisin-related proprotein
convertases furin, PC2, and PC3. Proc Natl Acad Sci USA 89:8822-8826.
21.Davidson HW, Wenzlau JM and OBrien RM. 2014. Zinc
transporter 8 (ZnT8) and beta cell function. Trends Endocrinol Metab. 25: 415424.
22.Kwok SC, Steiner DF, Rubenstein AH amd Tager HS. 1983.
Identification of a point mutation in the human insulin gene giving rise to a
structurally abnormal insulin (insulin Chicago).Diabetes.
32: 872-875.
23.Nishi M and Nanjo K. 2011. Insulin gene mutations and
diabetes. J Diabetes Investig. 2: 92100.
24.Bucchini D, Ripoche MA, Stinnakre MG, Desbois P, Lores
P, Monthioux E, Absil J, Lepesant JA, Pictet R and Jami J. 1986. Pancreatic
expression of human insulin gene in transgenic mice. Proc Natl Acad Sci U S A.
83: 25112515.
25.Howell SL. 1984.The mechanism of insulin secretion.
Diabetologia. 26: 319327.
26.Hay CW and Docherty K. 2006. Comparative analysis of
insulin gene promoters: implications for diabetes research. Diabetes. 55:
32013213.
27.Matsuoka TA, Artner I, Henderson E, Means A, Sander M
and Stein R. 2004. The MafA transcription factor appears to be responsible for
tissue-specific expression of insulin. Proc Natl Acad Sci U S A. 101:
29302933.
28.Vander Mierde D, Scheuner D, Quintens R, Patel R, Song
B, Tsukamoto K, Beullens M, Kaufman RJ, Bollen M and Schuit FC. 2007. Glucose
activates a protein phosphatase-1-mediated signaling pathway to enhance overall
translation in pancreatic beta-cells. Endocrinology. 148: 609617.
29.Mziaut H, Trajkovski M, Kersting S, Ehninger A,
Altkruger A, Lemaitre RP, Schmidt D, Saeger HD, Lee MS, Drechsel DN, Muller S
and Solimena M. 2006. Synergy of glucose and growth hormone signalling in islet
cells through ICA512 and STAT5. Nat Cell Biol. 8: 435445.
30.Itoh N and Okamoto H. 1980.Translational control of
proinsulin synthesis by glucose. Nature. 283: 100102.
31.Giddings SJ, Chirgwin J and Permutt MA. 1982. Effects
of glucose on proinsulin messenger RNA in rats in vivo. Diabetes. 31: 624629.
32.Wicksteed B, Uchizono Y, Alarcon C, McCuaig JF, Shalev
A and Rhodes CJ. 2007. A cis-element in the 5′ untranslated region of the
preproinsulin mRNA (ppIGE) is required for glucose regulation of proinsulin
translation. Cell Metab. 5: 221227.
33.Schmitz O, Rungby J, Edge L and Juhl CB. 2008. On
high-frequency insulin oscillations. Ageing Res Rev.7: 301305.
34.Suckale J and Solimena M. 2008. Pancreas islets in
metabolic signaling--focus on the beta-cell. Front Biosci.13: 71567171.
35.Nolan CJ and Prentki M.2008. The islet beta-cell: fuel responsiveand
vulnerable. Trends Endocrinol Metab 19: 285-291.
36.Nolan CJ, Madiraju MS,
Delghnngaro-Augusto V, Peyot ML, and Prentiki M.2006.Fatty acid signaling in the beta-cell and insulin
secretion. Diabetes; 55 (Suppl 2): S16-23.
37.Ruiz De Azua I, Gautam D,
Guettier JM and Wess J. 2011.
Novel insights into the function of beta-cell M3 muscarinic acetylcholine
receptors: Therapeutic implications. Trends Endocrinol Metab; 22: 74-80.
38.Ashcroft FM and Rorsman P.
2012. Diabetes mellitus and the beta cell: The last ten years. Cell 148:
1160-1171.
39.Zou
C-Y, Gong Y and Liang, J.2014. Metabolic
signaling of insulin secretion by pancreatic β-cell and its
derangement in type 2 diabetes. Eur. Rev. Med.
Pharmacol. Sci. 18: 2215-2227.
40.Sekine N, Cirulli V,
Regazzi R, Brown LJ, Gine E, Tamarit-Rodriguez J, Girotti M, Marie S, Macdonald
MJ, Wollheim CB and Rutter GA.1994. Low lactate dehydrogenase and high mitochondrial glycerol
phosphate dehydrogenase in pancreatic beta-cells. Potential role in nutrient
sensing. J Biol Chem. 269: 4895-4902.
41.Kwan EP and Gaisano HY.
2009. Rescuing the subprime meltdown in insulin exocytosis in diabetes.
Ann N Y Acad Sci. 1152: 154-164.
42.Nolan CJ, Damm P and Prentki
M. 2011. Type 2 diabetes
across generations: From pathophysiology to prevention and management. Lancet.
378: 169-181.
43.Wang Z and Thurmond DC. 2009. Mechanisms of biphasic insulin-granule
exocytosisroles of the cytoskeleton, small GTPases and SNARE proteins. J Cell
Sci. 122: 893-903.
44.Drucker DJ.
2006. The biology of incretin hormones. Cell Metab; 3: 153-165.
45.Ishii H, Sato Y, Takei M,
Nishio S and Komatsu M. 2011. Glucose-incretin interaction revisited. Endocrine J. 58:519-525.
46.Seino S, Takahashi H, Fujimoto W and Shibasaki T.
2009. Roles of cAMP signalling in insulin granule exocytosis. Diabetes Obes
Metab 11: 180-188.
47.Seino S and Shibasaki T. 2005. "PKA-dependent and
PKA-independent pathways for cAMP-regulated exocytosis". Physiol Rev. 85:
13031342.
48.Chang TW and Goldberg AL. 1978. The metabolic fates of
amino acids and the formation of glutamine in skeletal muscle. J Biol Chem.
253: 36853693.
49.Nauck MA, Bartels E, Orskov C, Ebert R and Creutzfeldt
W. 1993. Additive insulinotropic effects of exogenous synthetic human gastric
inhibitory polypeptide and glucagon-like peptide-1-(736) amide infused at
near-physiological insulinotropic hormone and glucose concentrations. JClin
Endocrinol Metab. 76: 912917.
50.Dixon G, Nolan J, McClenaghan N, Flatt PR and Newsholme
P. 2003. A comparative study of amino acid consumption by rat islet cells and
the clonal beta-cell line BRIN-BD11 - the functional significance of L-alanine.
J Endocrinol.179: 447454.
51.Itoh Y, Kawamata Y, Harada M, Kobayashi M, Fujii R,
Fukusumi S, Ogi K, Hosoya M, Tanaka Y, Uejima H, Tanaka H, Maruyama M, Satoh R,
Okubo S, Kizawa H, Komatsu H, Matsumura F, Noguchi Y, Shinohara T, Hinuma S,
Fujisawa Y and Fujino M. 2003. Free fatty acids regulate insulin secretion from
pancreatic beta cells through GPR40. Nature. 422: 173176.
52.Nadal
A, Rovira JM, Laribi O, Leon-quinto T, Andreu E, Ripoll C and Soria B. 1998. Rapid insulinotropic effect of 17beta-estradiol via a
plasma membrane receptor. FASEB J. 12: 13411348.
53.Peschke E, Peschke D, Hammer T and Csernus V. 1997.
Influence of melatonin and serotonin on glucose-stimulated insulin release from
perifused rat pancreatic islets in vitro. J Pineal Res. 23: 156163
54.Kulkarni RN, Wang ZL, Wang RM, Hurley JD, Smith DM,
Ghatei MA, Withers DJ, Gardiner JV, Bailey CJ and Bloom SR. 1997. Leptin
rapidly suppresses insulin release from insulinoma cells, rat and human islets
and, in vivo, in mice. J Clin Invest. 100: 27292736.
55.Guler HP, Schmid C, Zapf J and Froesch ER. 1989.
Effects of recombinant insulin-like growth factor I on insulin secretion and
renal function in normal human subjects. Proc Natl Acad Sci U S A. 86:
28682872.
56.Cañas
X, Fernández-López JA, Ardévol A, Adán C, Esteve M, Rafecas I, Remesar X and
Alemany M. 1995. Rat insulin
turnover in vivo. Endocrinology. 136: 3871-3876.
57.Sato H, Terasaki T, Mizuguchi H, Okumura K and Tsuji
A. 1991. Receptor-recycling model of clearance and distribution of insulin in
the perfused mouse liver. Diabetologia. 34: 613-621.
58.Duckworth WC, Bennett RG and Hamel FG. 1998. Insulin
degradation: progress and potential. Endocr Rev. 19: 608-624.
59.Rabkin R, Ryan MP and Duckworth WC.1984. The renal metabolism of insulin. Diabetologia.
27: 351-357.
60.Hamel FG, Peavy DE, Ryan MP and Duckworth WC. 1987. HPLC
analysis of insulin degradation products from isolated hepatocytes. Effects of
inhibitors suggest intracellular and extracellular pathways. Diabetes. 36: 702-708.
NOTA:Toda la información que se brinda en este artículo es de cará
cter investigativo y con fines académicos y de actualización para estudiantes y profesionales de la salud. En ningún caso es de carácter general ni sustituye el asesoramiento de un médico. Ante cualquier duda que pueda tener sobre su estado de salud, consulte con su médico o especialista.
Instituto de Medicina Tropical - Facultad de Medicina - Universidad Central de Venezuela.
Elaborado por el Centro de Análisis de Imágenes Biomédicas Computarizadas CAIBCO, caibco@ucv.ve
Este portal ha sido desarrollado gracias al apoyo del Fonacit