Micología
Características de las Conidias de Paracoccidioides brasiliensis
Generalidades sobre las conidias de P. brasiliensis
Estas estructuras fueron
observadas en aislamientos colombianos por Restrepo en 1970 (21), al realizar
un estudio encaminado a determinar el crecimiento de la fase miceliar de P.
brasiliensis en tierras; en aquel momento se anotó su semejanza
con las artroconidias intercalares y laterales, producidas por otros mohos patógenos
como Coccidioides immitis (21,22). Posteriormente, se describieron
con más detalle las características de tales propágulos
de P. brasiliensis (23). No fue posible, sin embargo, producirlas en
buen número por lo que se suspendió su estudio. A pesar de ello,
se mantuvo el interés por las conidias y años después (1985)
se emprenderían estudios metódicos encaminados a conocerlas mejor.
Se trabajó
entonces con varios aislamientos de P. brasiliensis provenientes de
pacientes con PCM, en un sistema de microcultivos preparados en medios pobres
(agar agua, dextrosa sales), en los cuales se intentó seguir, paso a
paso, el desarrollo y esporulación de los micelios. Se observó
así cómo en algunos aislamientos del hongo - y luego de 8 semanas
de incubación a 20-22º C - aparecían microconidias pedunculadas
y artroconidias tálicas intercalares, algunas de las cuales daban lugar
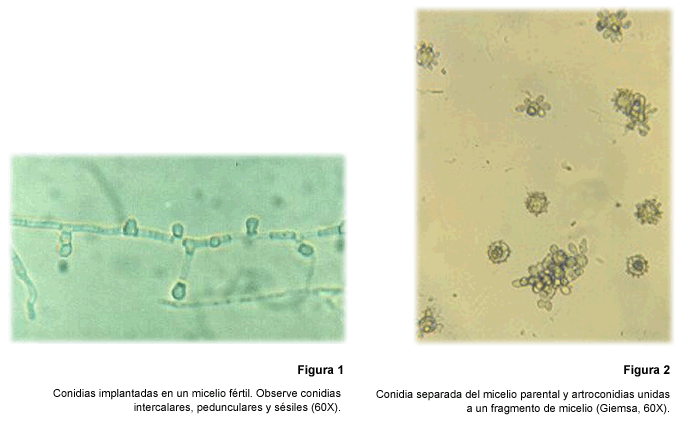
a un abultamiento lateral que resultaba en otra conidia (Fig. 1). Todas las
conidias eran pequeñas y medían menos de 5 µm; estas observaciones
permitieron comprobar las varias formas, el tamaño y la relación
con el micelio parental de las conidias (24).
A pesar del avance anterior, para
caracterizar las conidias era indispensable individualizarlas, separándolas
del micelio que las originaba. Después de múltiples intentos se
logró separarlas del micelio a partir de cultivos hechos en caja de Petri
utilizando los medios pobres en nutrientes ya mencionados e incubando por 2-3
meses a temperaturas menores a 20 º C (25) (Fig. 2). Bajo estas condiciones,
era posible obtener por caja de Petri hasta un millón de conidias, 80%
de las cuales se mostraban viables después de separadas (25).
En este momento, era posible trabajar
con las conidias como unidades independientes. Puesto que era necesario conocer
si poseían o no el dimorfismo térmico, se exploró la posible
influencia de la temperatura sobre las conidias. Para ello se retornó
a los microcultivos, sembrándolos con conidias e incubándolos
separadamente tanto a 20-22º C como a 36º C, haciendo observaciones
periódicas. A 20-22º C las conidias empezaban a emitir tubos germinales
antes de las 24 horas y a las 96 horas, presentaban ya micelios septados y ramificados.
Al incubar a 36º C las conidias adquirían inicialmente un aspecto
redondeado para dar lugar, 132 horas después, a las levaduras con gemación
múltiple (aproximadamente 40 µm diámetro) características
de P. brasiliensis (Fig. 3) (26). 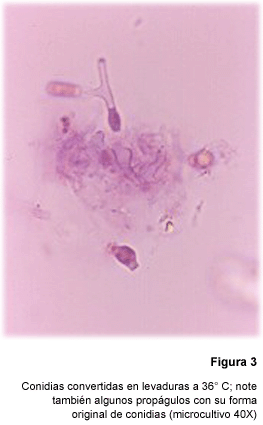
A pesar de su pequeño tamaño
(menos de 5 µm), las conidias presentaban el dimorfismo térmico
convirtiéndose, al elevar la temperatura a 36º C, en células
levaduras 9-10 veces mayores que ellas. Pero también eran igualmente
capaces de emitir micelios al ser incubadas a temperaturas más bajas
(20-22º C) (26). En otro contexto, los resultados anteriores revelaron
que la conidia de P. brasiliensis es una estructura inestable, de transición
y como tal, incapaz de mantenerse indefinidamente bajo esta forma; la influencia
de la temperatura determina si se hace micelio o levadura.
Como era conocido, ya que tanto el
micelio como las levaduras de P. brasiliensis eran multinucleadas (14),
se quiso averiguar si las conidias también lo eran. Se incubaron suspensiones
de conidias a 36º C por varias horas y se realizaron coloraciones especiales
con mitramicina para la observación de núcleos, haciendo la lectura
al microscopio de fluorescencia (27). Fue posible observar que las conidias
eran uninucleadas al momento de comenzar el experimento. Sin embargo, 72-96
horas después de su transición a levaduras, 80% de ellas presentaban
ya 4-5 núcleos (27). 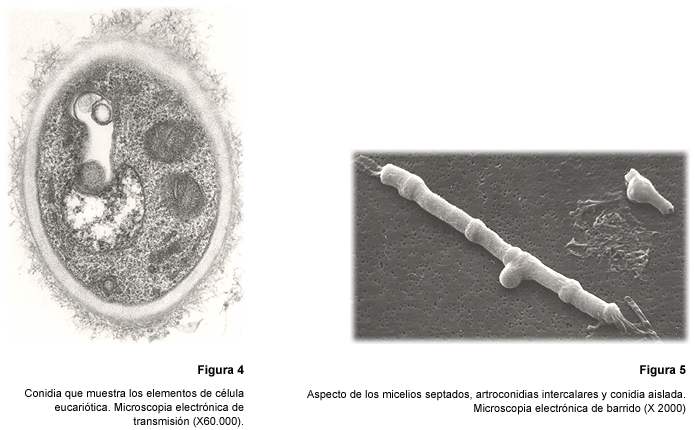
Una contribución
de la mayor importancia al conocimiento de la ultraestructura de las conidias
fue la realizada años más tarde (1991) por Samsonoff y Edwards
(28,29). Al utilizar microscopia electrónica, tanto de transmisión
como de barrido, fue posible demostrar la presencia de todos los componentes
esenciales de una célula eucariótica fisiológicamente competente;
a saber, un núcleo acompañado de nucleólo, mitocondrias,
ribosomas y peroxisomas, además de inclusiones de reserva, especialmente
aquellas compuestas por lípidos (Fig. 4). Adicionalmente, se observó
la presencia de abundante material fibrilar en la superficie celular (28,29).
Estos estudios permitieron configurar más precisamente el aspecto de
las artroconidias intercalares, de las conidias secundarias y de las pedunculadas
e igualmente, determinar la presencia de septos en los micelios del hongo (Fig.
5) y observar cómo muchos de aquellos aparecían vacíos
(28,29). Cuando se considera la escasa vitalidad de estos micelios, el tiempo
prolongado requerido para la producción de las conidias y el hecho de
que la producción de éstas se logre sólo bajo condiciones
de pobreza nutricional, es lógico señalar -como lo hiciera Edwards-,
que las conidias de P. brasiliensis representan un evento terminal
en la evolución de la fase miceliar del hongo (29). |